All animal housing and procedures have been approved by McGill University's Animal Care Committee.
Current research
Our lab works on three study systems, with a focus on spatial behaviour:

1) Behavioural and cognitive ecology of brood parasitism:
While most birds take care of their own young, some species like the brown-headed cowbird lay eggs in the nests of other species (obligate brood parasitism; see photo), while others like red-breasted mergansers lay eggs in the nests of conspecifics (facultative brood parasitism). This atypical behaviour involves several cognitive abilities to be successful. Our team is assessing the sexual preferences of cowbirds associated with potential mate cognitive abilities and the mergansers’ parasitic nest preferences according to genetic closeness between the nest host and parasitic individual.
Photo by Elsa Mardiné.
While most birds take care of their own young, some species like the brown-headed cowbird lay eggs in the nests of other species (obligate brood parasitism; see photo), while others like red-breasted mergansers lay eggs in the nests of conspecifics (facultative brood parasitism). This atypical behaviour involves several cognitive abilities to be successful. Our team is assessing the sexual preferences of cowbirds associated with potential mate cognitive abilities and the mergansers’ parasitic nest preferences according to genetic closeness between the nest host and parasitic individual.
Photo by Elsa Mardiné.

2) Spatial behaviour and cognition of ring-billed gull
With advanced GPS-tracking technology, we can now track free-ranging animals at a finer scale than ever before. We are examining whether the complexity of free-ranging movement in ring-billed gulls (see photo) is associated with cell-level variation in a part of the brain important for spatial memory, the hippocampus, and spatial cognition. We are also measuring diverse environmental contaminants (e.g., flame retardants, heavy metals) to determine whether contaminant load disrupts this behaviour-brain-cognition relationship.
Photo by Anna Lippold.
With advanced GPS-tracking technology, we can now track free-ranging animals at a finer scale than ever before. We are examining whether the complexity of free-ranging movement in ring-billed gulls (see photo) is associated with cell-level variation in a part of the brain important for spatial memory, the hippocampus, and spatial cognition. We are also measuring diverse environmental contaminants (e.g., flame retardants, heavy metals) to determine whether contaminant load disrupts this behaviour-brain-cognition relationship.
Photo by Anna Lippold.
3) European starling migration
Little is known on how the hippocampus changes in preparation for migration, a spatially demanding behaviour for many songbirds. In addition, exposure to pesticides can disrupt normal brain development. Our team is examining how exposure to pesticides (chlorpyrifos and imidacloprid) can affect neuron numbers and neurogenesis in the hippocampus (see photo) of European starlings and impact their migratory behaviour, from independence to fueling to peak migration.
Photo by Sereena Moore.
Little is known on how the hippocampus changes in preparation for migration, a spatially demanding behaviour for many songbirds. In addition, exposure to pesticides can disrupt normal brain development. Our team is examining how exposure to pesticides (chlorpyrifos and imidacloprid) can affect neuron numbers and neurogenesis in the hippocampus (see photo) of European starlings and impact their migratory behaviour, from independence to fueling to peak migration.
Photo by Sereena Moore.
Work prior to Tenure track position
1) Cognition of avian brood parasites
Summary: My co-authors and I measured sex differences in spatial memory in brown-headed cowbirds (Molothrus ater) using different tasks. Female cowbirds outperformed males on a task that resembled their behaviour in the wild.
Brood parasites lay their eggs in the nests of other species of birds. In some brood parasites, such as brown-headed cowbirds (Molothrus ater), females are the only member of the pair to search for, parasitize, and re-visit host nests. As such, females are expected to have better spatial memory than males. My co-authors and I showed that female cowbirds outperformed males on a spatial memory food cup task that resembled host nest visits in the wild in which subjects needed to move through space and remember a specific location for 24 hours (Guigueno et al. 2014, Biology Letters). However, females did not outperform males on a stationary touchscreen task in which subjects only had to remember a location on a screen for up to 60 sec. In fact, males outperformed females on this task, suggesting there may be trade-offs between different forms of spatial memory (Guigueno et al. 2015, Plos One). We also describe our work on cognition in brood parasites in an upcoming book chapter, Hippocampus and Spatial Memory in Brood Parasitic Cowbirds to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).
Summary: My co-authors and I measured sex differences in spatial memory in brown-headed cowbirds (Molothrus ater) using different tasks. Female cowbirds outperformed males on a task that resembled their behaviour in the wild.
Brood parasites lay their eggs in the nests of other species of birds. In some brood parasites, such as brown-headed cowbirds (Molothrus ater), females are the only member of the pair to search for, parasitize, and re-visit host nests. As such, females are expected to have better spatial memory than males. My co-authors and I showed that female cowbirds outperformed males on a spatial memory food cup task that resembled host nest visits in the wild in which subjects needed to move through space and remember a specific location for 24 hours (Guigueno et al. 2014, Biology Letters). However, females did not outperform males on a stationary touchscreen task in which subjects only had to remember a location on a screen for up to 60 sec. In fact, males outperformed females on this task, suggesting there may be trade-offs between different forms of spatial memory (Guigueno et al. 2015, Plos One). We also describe our work on cognition in brood parasites in an upcoming book chapter, Hippocampus and Spatial Memory in Brood Parasitic Cowbirds to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).




Images of food cup apparatus (two left images above) and touchscreen operant conditioning chamber (two right images above).
2) The brain of avian brood parasites
Summary: We measured the volumes and levels of neurogenesis (ie., the production of new neurons) of the hippocampus (region responsible for spatial memory) and of two song control nuclei (regions responsible for song learning and production) in cowbirds and a non-parasitic relative, the red-winged blackbird (Agelaius phoeniceus). Female cowbirds had higher levels of hippocampal neurogenesis than males, with no sex difference in red-winged blackbirds. Sex, species, and seasonal changes in the volume and levels of neurogenesis in the song control nuclei of cowbirds and red-winged blackbirds varied with song production.
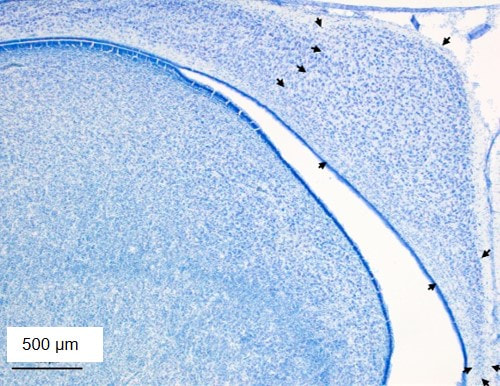
Hippocampus - The region of the brain responsible for spatial memory is the hippocampus. Previous research on North and South American brood parasites have shown that in species in which females search for host nests and males do not, females have a larger hippocampus than males, with no sex difference in species in which both sexes search and in non-brood-parasitic species. We found that in both brown-headed cowbirds and the non-brood-parasitic relative red-winged blackbird (Agelaius phoeniceus), females had a larger hippocampus than males (Guigueno et al 2016b, Developmental Neurobiology), in contrast with previous research. However, we also measured neurogenesis in the hippocampus. Neurogenesis is the production of new neurons. We found that female cowbirds had significantly more hippocampal neurogenesis than male cowbirds, with no sex difference in red-winged blackbirds (Guigueno et al. 2016b). In addition, we found that in cowbirds, hippocampal neurogenesis was significantly greater in post-breeding condition than in breeding condition, indicating that neurogenesis may play an important role in forgetting past memories (i.e., locations of host nests from the previous breeding season) to facilitate the acquisition of new memories (i.e., locations of host nests in the upcoming breeding season) (Guigueno et al. 2016b). We also describe our work on the hippocampus of brood parasites in an upcoming book chapter, Hippocampus and Spatial Memory in Brood Parasitic Cowbirds to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).
2) The brain of avian brood parasites
Summary: We measured the volumes and levels of neurogenesis (ie., the production of new neurons) of the hippocampus (region responsible for spatial memory) and of two song control nuclei (regions responsible for song learning and production) in cowbirds and a non-parasitic relative, the red-winged blackbird (Agelaius phoeniceus). Female cowbirds had higher levels of hippocampal neurogenesis than males, with no sex difference in red-winged blackbirds. Sex, species, and seasonal changes in the volume and levels of neurogenesis in the song control nuclei of cowbirds and red-winged blackbirds varied with song production.
Hippocampus - The region of the brain responsible for spatial memory is the hippocampus. Previous research on North and South American brood parasites have shown that in species in which females search for host nests and males do not, females have a larger hippocampus than males, with no sex difference in species in which both sexes search and in non-brood-parasitic species. We found that in both brown-headed cowbirds and the non-brood-parasitic relative red-winged blackbird (Agelaius phoeniceus), females had a larger hippocampus than males (Guigueno et al 2016b, Developmental Neurobiology), in contrast with previous research. However, we also measured neurogenesis in the hippocampus. Neurogenesis is the production of new neurons. We found that female cowbirds had significantly more hippocampal neurogenesis than male cowbirds, with no sex difference in red-winged blackbirds (Guigueno et al. 2016b). In addition, we found that in cowbirds, hippocampal neurogenesis was significantly greater in post-breeding condition than in breeding condition, indicating that neurogenesis may play an important role in forgetting past memories (i.e., locations of host nests from the previous breeding season) to facilitate the acquisition of new memories (i.e., locations of host nests in the upcoming breeding season) (Guigueno et al. 2016b). We also describe our work on the hippocampus of brood parasites in an upcoming book chapter, Hippocampus and Spatial Memory in Brood Parasitic Cowbirds to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).
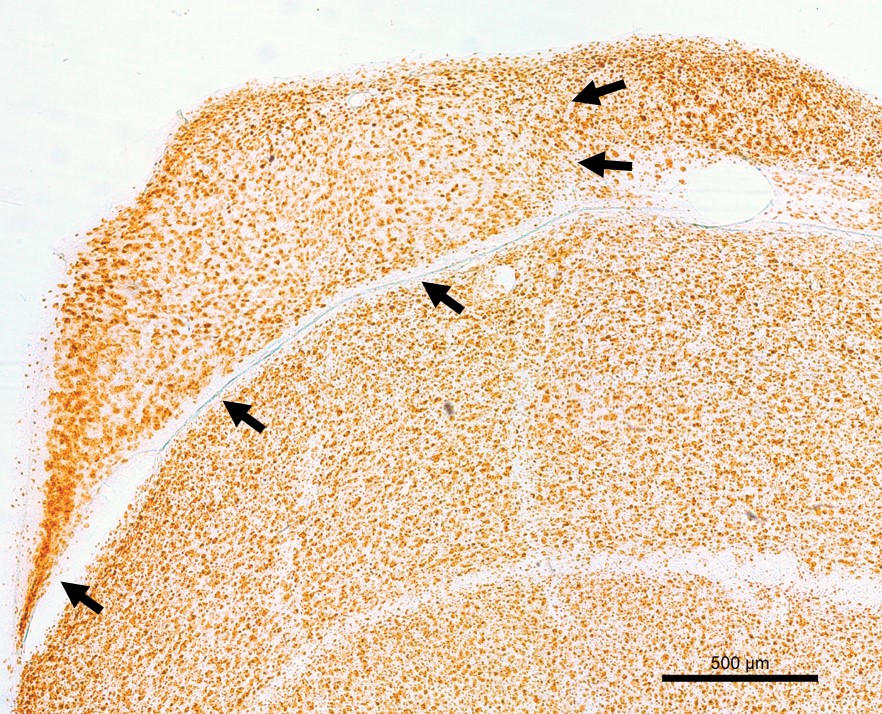

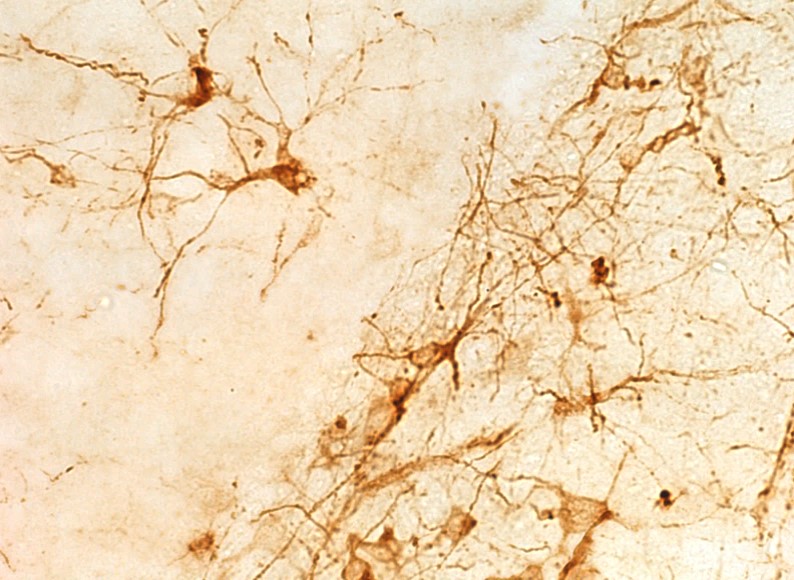
Images above are coronal sections of the cowbird hippocampus. Image to the left is a section labeled via NeuN immunohistochemistry, with all mature neurons labeled. Arrows indicate the boundaries of the hippocampus. Two other images are of doublecortin (DCX) immunohistochemistry. Doublecortin is a protein only present in immature neurons. Migrating immature neurons are called fusiform cells (bottom right arrow in middle image) whereas immature neurons that have reached their final destination are called round cells (top left arrow in middle image). The doublecortin image to the right is an example of immunoreactivity in the subventricular zone (ventral side of the hippocampus), where new neurons are born.
Song control nuclei - We measured sex, seasonal, and species differences in two song control nuclei in cowbirds and blackbirds; HVC and the robust nucleus of the arcopalllium (RA). HVC and RA play play important roles in song learning and song production, two components that differ between cowbirds and blackbirds. Blackbirds learn their song while still in the nest and cowbirds, because they are brood parasites, learn their song later in life (i.e., late summer of their hatch year) when they interact with members of their own species. In addition, female cowbirds do not sing at all whereas female blackbirds do. My co-authors and I found that in breeding condition, female blackbirds had a more discernible HVC and a larger RA than female cowbirds (Guigueno et al. 2016a, Developmental Neurobiology). Females of both species had a smaller HVC and RA than males, with more neurogenesis in the HVC. Finally, males of both species had a smaller HVC with higher levels of neurogenesis in post-breeding condition when song is more plastic (i.e., variable) (Guigueno et al. 2016a).
Song control nuclei - We measured sex, seasonal, and species differences in two song control nuclei in cowbirds and blackbirds; HVC and the robust nucleus of the arcopalllium (RA). HVC and RA play play important roles in song learning and song production, two components that differ between cowbirds and blackbirds. Blackbirds learn their song while still in the nest and cowbirds, because they are brood parasites, learn their song later in life (i.e., late summer of their hatch year) when they interact with members of their own species. In addition, female cowbirds do not sing at all whereas female blackbirds do. My co-authors and I found that in breeding condition, female blackbirds had a more discernible HVC and a larger RA than female cowbirds (Guigueno et al. 2016a, Developmental Neurobiology). Females of both species had a smaller HVC and RA than males, with more neurogenesis in the HVC. Finally, males of both species had a smaller HVC with higher levels of neurogenesis in post-breeding condition when song is more plastic (i.e., variable) (Guigueno et al. 2016a).
|
|
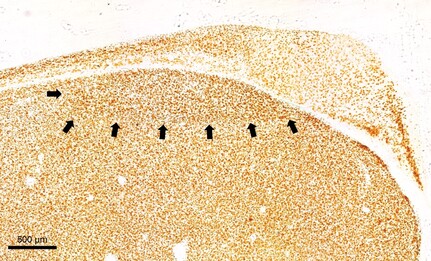
Images of HVC (left) and robust nucleus of the arcopallium (RA; right), with mature neurons immunolabeled via NeuN immunohistochemistry.
3) Cues used in decision making in a cowbird host
Summary: My co-author and I tested four cues that could be used by yellow warblers to reject their clutches parasitized by cowbirds: adding a model cowbird egg to their nest at sunrise, removing a warbler egg later in the morning, placing a model of a laying female cowbird on the nest before sunrise, and placing a robotic egg-removing cowbird at the nest later in the morning. We found that warblers used the model cowbird egg and the egg-removing cowbird robot as cues to reject their parasitized clutch, producing rejection frequencies similar to those recorded at naturally parasitized nests.
Some hosts of brown-headed cowbirds reject the parasitic egg from their nest. Hosts can reject an egg by grasping it between their mandibles and removing it (grasp-ejection), puncturing the egg with their bill (puncture-ejection), or deserting the nest and nest site. There is one other method of rejection that is less common: egg burial. Egg burial involves the hosts burying the cowbird egg, usually along with its own eggs, and building a new superimposed nest on the previous nest and eggs. Yellow warblers are one of very few hosts that consistently uses this method of rejection. Only about 50% of female warblers naturally parasitized by cowbirds were known to reject cowbird eggs (primarily by burial) at our field site at Delta Marsh, Manitoba. However, nests that were experimentally parasitized by researchers (i.e., egg added to host nest by hand) produced a much lower rejection frequency. My co-authors and I built a robot from a taxidermied specimen of a female brown-headed cowbird, which I used to simulate a more realistic simulation of a host nest visit by the parasite (Guigueno and Sealy 2011, Animal Behaviour). Cowbirds will return to some nests they have parasitized to remove a host egg. Our female robot had an artificial warbler egg in its bill and a simulation of a nest visit consisted of the robot having its head in the nest cavity and the head moving up from the cavity. Warblers that were aggressive toward our robot were more likely to subsequently reject their experimentally parasitized clutch, indicating that interaction with the parasite is a critical component of decision-making in parasitized hosts (Guigueno and Sealy 2011). I published seven other papers examining egg rejection in hosts of brood parasites (see publications), some of which are related to the theme described below (nest sanitation).
Summary: My co-author and I tested four cues that could be used by yellow warblers to reject their clutches parasitized by cowbirds: adding a model cowbird egg to their nest at sunrise, removing a warbler egg later in the morning, placing a model of a laying female cowbird on the nest before sunrise, and placing a robotic egg-removing cowbird at the nest later in the morning. We found that warblers used the model cowbird egg and the egg-removing cowbird robot as cues to reject their parasitized clutch, producing rejection frequencies similar to those recorded at naturally parasitized nests.
Some hosts of brown-headed cowbirds reject the parasitic egg from their nest. Hosts can reject an egg by grasping it between their mandibles and removing it (grasp-ejection), puncturing the egg with their bill (puncture-ejection), or deserting the nest and nest site. There is one other method of rejection that is less common: egg burial. Egg burial involves the hosts burying the cowbird egg, usually along with its own eggs, and building a new superimposed nest on the previous nest and eggs. Yellow warblers are one of very few hosts that consistently uses this method of rejection. Only about 50% of female warblers naturally parasitized by cowbirds were known to reject cowbird eggs (primarily by burial) at our field site at Delta Marsh, Manitoba. However, nests that were experimentally parasitized by researchers (i.e., egg added to host nest by hand) produced a much lower rejection frequency. My co-authors and I built a robot from a taxidermied specimen of a female brown-headed cowbird, which I used to simulate a more realistic simulation of a host nest visit by the parasite (Guigueno and Sealy 2011, Animal Behaviour). Cowbirds will return to some nests they have parasitized to remove a host egg. Our female robot had an artificial warbler egg in its bill and a simulation of a nest visit consisted of the robot having its head in the nest cavity and the head moving up from the cavity. Warblers that were aggressive toward our robot were more likely to subsequently reject their experimentally parasitized clutch, indicating that interaction with the parasite is a critical component of decision-making in parasitized hosts (Guigueno and Sealy 2011). I published seven other papers examining egg rejection in hosts of brood parasites (see publications), some of which are related to the theme described below (nest sanitation).



We examined whether the following cues elicited egg rejection in yellow warblers: a model of a laying female cowbird (left image), a model cowbird egg in the nest (middle image), the removal of a warbler egg (no image), and/or an egg-removing cowbird with an artificial warbler egg in its bill.
4) The importance of nest sanitation in the evolution of egg rejection behaviour in hosts of brood parasites
Summary: Nest sanitation behaviour may be a pre-adaptation for the evolution of egg rejection behaviour in hosts of brood parasites.
The motor patterns involved in removing a parasitic egg from the nest are similar to those involved in removing debris from the nest to keep it clean. My most-cited paper, Guigueno & Sealy (2012; Journal of Ornithology) is a review stemming from an undergraduate class project that has played a pivotal role in stimulating new research on nest sanitation. In our review, my co-author and I brought together evidence showing that nest sanitation is ubiquitous among songbirds and may be a pre-adaptation for egg rejection in hosts of brood parasites, thus also making a landmark contribution to the field of avian brood parasitism. I also published my Honours project in which I tested the influence of shape and volume on the rejection behaviour of yellow warblers to determine whether more debris-shaped objects were more likely to be rejected (see images below; Guigueno and Sealy 2009, Ethology). We also describe our work on nest sanitation in an upcoming book chapter, Implications of Nest Sanitation in the Evolution of Egg Rejection In Avian Brood Parasitism to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).
4) The importance of nest sanitation in the evolution of egg rejection behaviour in hosts of brood parasites
Summary: Nest sanitation behaviour may be a pre-adaptation for the evolution of egg rejection behaviour in hosts of brood parasites.
The motor patterns involved in removing a parasitic egg from the nest are similar to those involved in removing debris from the nest to keep it clean. My most-cited paper, Guigueno & Sealy (2012; Journal of Ornithology) is a review stemming from an undergraduate class project that has played a pivotal role in stimulating new research on nest sanitation. In our review, my co-author and I brought together evidence showing that nest sanitation is ubiquitous among songbirds and may be a pre-adaptation for egg rejection in hosts of brood parasites, thus also making a landmark contribution to the field of avian brood parasitism. I also published my Honours project in which I tested the influence of shape and volume on the rejection behaviour of yellow warblers to determine whether more debris-shaped objects were more likely to be rejected (see images below; Guigueno and Sealy 2009, Ethology). We also describe our work on nest sanitation in an upcoming book chapter, Implications of Nest Sanitation in the Evolution of Egg Rejection In Avian Brood Parasitism to be published in Avian Brood Parasitism - Behaviour, Ecology, Evolution and Coevolution (Edited by M. Soler).



Rejection responses of non-egg-shaped objects added to the nests of yellow warblers. Left: dumbbell-shaped object (two smaller spheres glued together) buried along with two warbler eggs, with a new clutch of four eggs on top. Middle: star-shaped object selectively buried within the lining of the nest. Right: female yellow warbler grasp-ejecting a star-shaped object. Warblers normally do not eject cowbird eggs, likely because they are too large for them to be grasped.
5) Social learning in Trinidadian guppies (Poecilia reticulata)
Currently as part of my postdoctoral research with Simon Reader, I am examining the effects of stress on social learning and the brain in this highly social fish. With the cowbird research described above, I focused on a specialized form of cognition, spatial memory, and its neural basis, the hippocampus. In contrast, social learning is a cognitive trait that is linked to general intelligence.
A MSc student and I recently validated an a new phospho-S6 antibody for immunohistochemistry in guppies, which is a marker of brain activity (see photo to the right below; photo to the left by Harald Olsen). I plan to share results from this brain study and the social learning study at upcoming conferences.
5) Social learning in Trinidadian guppies (Poecilia reticulata)
Currently as part of my postdoctoral research with Simon Reader, I am examining the effects of stress on social learning and the brain in this highly social fish. With the cowbird research described above, I focused on a specialized form of cognition, spatial memory, and its neural basis, the hippocampus. In contrast, social learning is a cognitive trait that is linked to general intelligence.
A MSc student and I recently validated an a new phospho-S6 antibody for immunohistochemistry in guppies, which is a marker of brain activity (see photo to the right below; photo to the left by Harald Olsen). I plan to share results from this brain study and the social learning study at upcoming conferences.

|
|
6) Ecotoxicology; linking chemical contamination to changes in behaviour and the brain
Summary: My co-authors and I examined the effect of mercury and flame retardants on neuroanatomy (volume of targeted brain regions) and behaviour, alongside metabolic rate, hormones, and gene expression.
Last, but certainly not least, I have incorporated my skills in animal behaviour and neuroscience to a more applied context, ecotoxicology. I have been writing ecotoxicology manuscripts as additional projects since my Master's degree (Guigueno et al. 2012, Environment International; Elliott et al. 2012, Environmental Science and Technology; Miller et al. 2014, Environmental Pollution; Miller et al. 2015, Science of the Total Environment) and now, as a post-doctoral fellow, my co-authors and I have looked at the effects of mercury on the volume of song control nuclei in zebra finches (Taeniopygia guttata) (Yu et al. 2017, Neurotoxicology) and flame retardants on the volume of the hippocampus in American kestrel (Falco sparverius) hatchlings (image below; Guigueno et al. 2018, Guigueno et al. In Press). In addition, I am currently examining the effects of a currently used flame retardant and plasticizer, triphenyl phosphate (TPHP), on behaviour, hormones, metabolism, and gene expression in Japanese quail (Coturnix japonica) chicks (images below). Changes in the brain and behaviour due to chemical contamination are ecologically relevant as they can produce effects at the level of the population.
6) Ecotoxicology; linking chemical contamination to changes in behaviour and the brain
Summary: My co-authors and I examined the effect of mercury and flame retardants on neuroanatomy (volume of targeted brain regions) and behaviour, alongside metabolic rate, hormones, and gene expression.
Last, but certainly not least, I have incorporated my skills in animal behaviour and neuroscience to a more applied context, ecotoxicology. I have been writing ecotoxicology manuscripts as additional projects since my Master's degree (Guigueno et al. 2012, Environment International; Elliott et al. 2012, Environmental Science and Technology; Miller et al. 2014, Environmental Pollution; Miller et al. 2015, Science of the Total Environment) and now, as a post-doctoral fellow, my co-authors and I have looked at the effects of mercury on the volume of song control nuclei in zebra finches (Taeniopygia guttata) (Yu et al. 2017, Neurotoxicology) and flame retardants on the volume of the hippocampus in American kestrel (Falco sparverius) hatchlings (image below; Guigueno et al. 2018, Guigueno et al. In Press). In addition, I am currently examining the effects of a currently used flame retardant and plasticizer, triphenyl phosphate (TPHP), on behaviour, hormones, metabolism, and gene expression in Japanese quail (Coturnix japonica) chicks (images below). Changes in the brain and behaviour due to chemical contamination are ecologically relevant as they can produce effects at the level of the population.


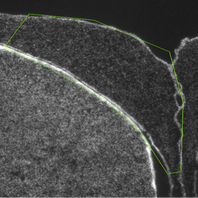
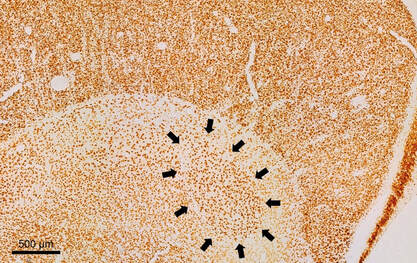
Multiple endpoints measured in the context of embryonic flame retardant exposure in birds. Left: Coronal section of the hippocampus of a hatchling American kestrel. Boundaries are indicated by arrows. All cells were labeled using a Nissl stain. Middle: Japanese quail chicks labeled with laboratory tape for behavioural analyses. Right: A Japanese quail chick in a respirometry chamber to measure its resting metabolic rate.